はじめに
背の高い茎のロゼット (巨大なロゼットとも呼ばれる) は、南アメリカのアンデス北部、東アフリカの高地、ハワイ諸島とカナリア諸島の大きな火山など、多くの熱帯の山々の標高の高い場所で育ちます。樹木とは対照的に、これらの植物の木質の幹は年輪を生成しないため、年齢を簡単に決定することはできません。代わりに、茎の高さの変化は、植物の年齢を推定するためのプロキシとして日常的に使用されてきました。過去 50 年間に、いくつかの茎ロゼット種の伸長率が測定されました。小さな集団または茎ロゼットのグループについて決定された平均成長率は、植物の年齢を計算するために使用されています。これらの植物が生育する山岳地帯へのアクセスが困難であり、さまざまな物流上の理由により、ほとんどの生育研究は短期間、多くの場合 1 年未満から数年に制限されています。
アンデスの茎のロゼットは、植物が修道士に似ていると言われているため、南アメリカでは frailejones (単数:frailejón、「大きな修道士」) として集合的に知られています。 frailejones は、エクアドル中央部からベネズエラ北部まで、1200 km 以上の距離にわたってアンデス山脈に沿って見られます (Cuatrecasas, 2013)。ここで報告された研究は、北緯 8 度 52 分、ベネズエラ、シエラ ラ クラタのパラモ ピエドラス ブランカスにある、背が高く豊富な 2 種類の frailejón 種に焦点を当てています。伸長率は、76 Coespeletia timotensis について測定されました。 および 38 Espeletia schultzii 標高 4270 ~ 4525 メートル (~14010 ~ 14850 フィート) で、10 ~ 20 年の範囲の期間。植物は繰り返し撮影の助けを借りて移動しました。この山脈では、C.チモテンシス 近くのシエラネバダデメリダでは4780m(〜15685 ')で発見されますが、〜4565 m(〜14980 ')で最高高度に達します。実際、この植物は、アンデスの 144 の frailejón 種のいずれかで報告されている最高の標高で生育します (Diazgranados, 2012)。 E.シュルツィー 、研究された他の植物も、この範囲で最大 4300 メートル (~14110’) まで一般的です。
茎ロゼットの気候的背景と成長
高地の過酷な気候条件は、成長の遅いフレレジョーンズを含むすべての植物の成長を著しく制限します。氷点下の気温がほぼ毎晩発生し、多くの地面に霜が降り、大部分の苗木が枯死します。永久凍土層の下限 - 平均土壌温度が 0°C を下回る - は、ピエドラス ブランカスの 4550 ~ 4600 m で発生します (Schubert, 1975)。極度の土壌干ばつも 4 か月にわたる乾期に発生し、植物にさらにストレスを与えます。アンデス北部では、植物の上限は、熱不足と生理的乾燥によって引き起こされる寒さと乾燥の限界です。困難な高山の条件にもかかわらず、一部のフレレジョンは驚異的なサイズに成長します。 C.チモテンシス 多くの場合、高さは約 3.5 m に達します (写真)。 E.シュルツィー はるかに短く、身長は 1.5 m 以下です。


これらの写真は、C のグループを示しています。チモテンシス 〜4385メートル。右側のものは、ピエドラス ブランカスで見つけた中で最も高いロゼットの 1 つです。 1982年1月10日、最上葉先端までの高さは291±2cm(写真a)。 2000 年 2 月 29 日、18 年 2 か月の期間の後、21 cm から 312±2 cm に成長しました (写真 b)。約 1.16 cm/年の比成長率 (以下の説明を参照) を使用すると、この植物の推定年齢は約 269 歳になります。もちろん、これはおそらく誤って、この植物の寿命の間、均一な成長率を前提としています.左から 2 番目に高いロゼットは、同じ期間にさらに成長したようです (~28 cm、~1.54 cm/年)。スケールの長さは 180 cm です。
短い期間にわたる以前の調査では、成長の遅い個体間で拡大率のかなりの変動性を検出することはできませんでしたが、10年から20年にわたって伸びを評価することで、小さな変動を最大化することができ、植物間で観察された成長差は〜34cm。 C の以前の平均成長率。チモテンシス 個体群は 1.5 cm/年、E では 1.0 cm/年と推定されました。シュルツィー (スミス、1981)。この長期研究中の平均胴体伸長率は有意差はなく、それぞれ 1.24 ± 0.46 cm/年、および 1.04 ± 0.40 cm/年でした。しかし、驚くべき全く予想外の結果の 1 つは、これらの平均率が、いずれかの種の個々の植物間の広範な変動を隠していたことです。
植物の生長の変動の考えられる原因
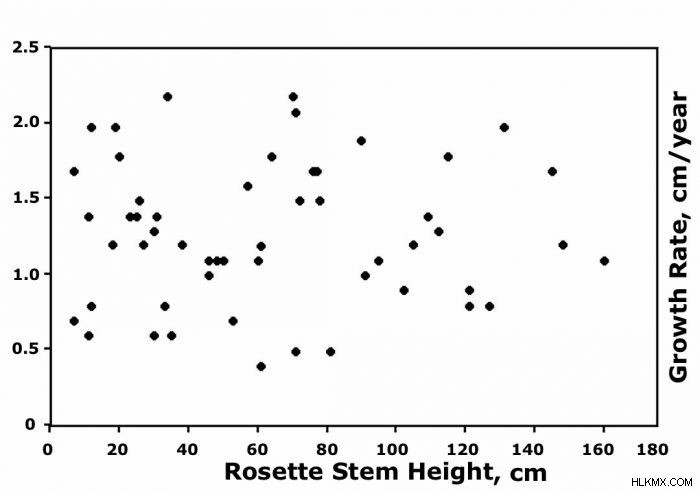
互いに近くに生育し、同様の地形的位置と土壌タイプを占め、同じ側面を持つ植物でさえ、極端な変動性を示しました.グラフは、生存している 60 匹の C に隣接する 10 年間、約 2 か月間の茎の成長率 (cm/年) を示しています。チモテンシス 〜4285 mの谷底に沿って〜0.4〜2.5 cm /年の間で変動しました。残りの 31 個の E の比較可能な散布図。シュルツィー は、成長率が年間約 0.4 ~ 2.4 cm の範囲で変動することを示しました。重要なことに、このグラフは初期の植物サイズが平均成長と関連していなかったことも示しており、これはランダムであり、植物の年齢とは無関係であるように思われました.
おそらく、個々の植物間の違いは、物理的なマイクロサイトの特徴の違い (短距離で変化する可能性があります)、または特定の植物の近くの生物学的条件のいずれかによって引き起こされる可能性があります。フレレホンの成長に影響を与えることが知られているマイクロサイトの要因には、土壌水分含有量、土壌栄養素のレベル、近くの岩の存在、および隣接する植物間の競合的相互作用が含まれます.
植物は、個々の茎ロゼットの成長に 2 つの方法で影響を与える可能性があります。 Frailejón の開発は、ナース クッション植物または岩の隣で発芽させることによって促進できます。どちらも霜の頻度と障害を減らし、一般的に土壌の水分状態を改善します。他の相互作用は、競争を通じて成長を阻害する可能性があります。たとえば、砂漠の柱状サボテンの研究では、植物のすぐ近くにある植生の豊富さが、成長率に悪影響を及ぼす「干渉要因」を生み出すことがわかっています。この研究では競合は分析されませんでしたが、パラモ植生に関する以前のデータは、C.チモテンシス スタンドには豊富な植生があり、64% の植物被覆と 37 の維管束植物種がありました。 E.シュルツィー 林分も植生が密集しており、45 の植物種と 61.5% の植物被覆がありました。
死亡による植物の消耗
研究期間の終わりまでに十分な数の植物が生き残り、重要な統計分析を実行できましたが、植物の死亡率により、両方の脆弱な個体群が約 18 ~ 19% 減少しました。 C.チモテンシス 実生はおそらく、凍上および/または半野生の牛による閲覧によって殺されました。背が高く、成熟した C.チモテンシス この種の一般的なイベントである植物の倒壊によって間引きされました。これはおそらく、霜の乱れによる斜面の動きと地面の移動に関連していました。土壌の不安定性は背の高いセネシオ ケニオデンドロンにも影響します ケニア山 (アフリカ) の茎ロゼットでは、傾斜した「酔っ払いスタンド」が急で乱れた斜面に発生します (Mahaney, 1980)。かなりの葉の軟化と樹脂状化合物の含有量が高いため、牛は E.E. を食べません。シュルツィー;また、おそらく身長が低いため、これらの frailejones は傾いてもほとんど影響を受けず、ほとんどの枯れた植物は倒れずに立ったままでした.
結論
結論として、茎とロゼットの成長率は、隣接する植物であっても、平均を使用して 6 ~ 16 倍変化します。 現在までの率個人 植物は無意味に思えます。特定の植物の特定の成長データが不足しているため、このアプローチは賢明ではないようであり、将来の研究では中止する必要があります.いずれにせよ、パラモ フライレジョーンズの正確な年代測定を重視することは、二次的なものかもしれません。これらの長命の植物の最も注目すべき点は、アンデス高地の敵対的な環境にいかに驚くほど適応しているか、そしてどのように進化して幅広い山の生息地を占めるようになったかということです。 C.チモテンシス 他のどのアンデス フライレホンよりも高い標高で生育するこの植物は、アンデス高地植物の最も顕著な進化の頂点を占める可能性があります (Cuatrecasas、2013)。
参考文献:
<オール>Memoirs New York Botanical Garden, 107, 1-689.
Phytokeys, 16, 1-52. https://phytokeys.pensoft.net/articles.php?id=1436. www.phytokeys.com
25、492-497。
アンデス。 Eiszeitalter und Gegenwart, 26, 196-211.